
domingo, 15 de fevereiro de 2009
Qual assunto na área de Zoologia você gostaria de ver aqui no ZOOBIOLOGY?
Bem caro visitante, você tambem é um elo pensante neste blog!. Fique avontade para escolher os proximos assuntos do Universo Zoológico.
sexta-feira, 13 de fevereiro de 2009
ZOOBIOLOGY
Segundo o grande Catedrático George Bartholomew "Se alguém aquacionar a ignorância como razão entre o que se sabe e o que está disponível para o conhecido...cada investigador biológico fica mais ignorante a cada dia que passa". E é com o propósito primordial deste blog tentar complementar minimamente a lacuna informativa no universo biológico.
Autor
Carlos Rodrigues
Autor
Carlos Rodrigues
Assinar:
Postagens (Atom)


Charles Robert Darwin (1809-1882)
ENSAIO
Nascido no século 19, Charles Robert Darwin (1809-1882) é o cientista mais conhecido e reconhecido no século 21, superando em notoriedade gigantes como o físico inglês Isaac Newton (1643-1727), o filósofo alemão Karl Marx (1818-1883), o neurologista austría-co Sigmund Freud (1856-1939) e o físico alemão Albert Einstein (1879-1955). Darwin foi o criador da teoria evolutiva que explica a origem de todas as espécies por meio da seleção natural. Essa teoria, após ter sido elaborada por ele durante décadas, foi lançada a público há exatos 150 anos, com a leitura, em sessão da Royal Society, principal academia de ciências da Inglater-ra, de cartas do próprio Darwin e do também inglês Alfred Wallace (1823-1913), que chegou bem mais tarde a conclusões semelhantes. Embora tenha mudado radicalmente a forma como o homem percebia a natureza, a teoria da evolução dificilmente explica o enorme prestígio que Darwin tem ainda hoje.
Na verdade, a contribuição de Darwin para o desenvolvimento do conhecimento humano vai muito além da teoria evolutiva: ele foi pioneiro na geologia, criando teorias importantes para o surgimento de ilhas oceânicas e explicações corretas para os mecanismos que geram os movimentos ascendentes da cordilheira dos Andes. Para o biólogo austríaco Konrad Lorenz (1903-1989), Darwin foi ainda o iniciador da etologia, ciência que estuda o comportamento dos animais. Além disso, pode ser considerado o mentor da moderna ecologia, por ter criado os conceitos de ‘nicho ecológico’ e ‘ecossistema’, ao afirmar que cada espécie ocupa um lugar determinado (nicho) na “economia da natureza” (ecossistema). Nessa mesma área, Darwin demonstrou a importância funda-mental, para o funcionamento dos ecossistemas, de relações como competição, predação e mutualismo entre os seres vivos e interações entre estes e o ambiente. Na taxonomia, a rainha das ciências biológicas, ele fez contribuições notáveis, criando os conceitos de ‘espécies em estado nascendi’ e de ‘espécie fóssil’. Naturalista, biólogo, etólogo, taxonomista, geólogo, ecó-logo e, como se fosse pouco, pai da biogeografia. Suas contribuições fundamentais em todas essas ciências podem explicar por que Dar-win é um gigante entre gigantes.
De avô para neto
O criador da teoria da evolução nasceu em 1809 em uma família rica, com tradição na medicina. O pai, Robert Darwin (1766-1848), era médico, e o avô paterno, Erasmus Darwin (1731-1802) era, além de médico, um poeta e um adepto das idéias sobre evolução do biólogo francês Jean-Baptiste Lamarck (1744-1829), que acreditava na transmutação das espécies e criou uma teoria para explicá-la. Erasmus escreveu Zoonomia, um vo-lumoso livro sobre o tema da ori-gem das espécies, lido e apreciado pelo neto. Aos oito anos, Charles Darwin perdeu a mãe e entrou pa-ra a escola. Os oito anos passados nessa escola foram considerados pelo próprio Darwin uma grande e enfadonha perda de tempo. Seu baixo desempenho fez com que seu pai o enviasse para a Universidade de Edimburgo, na Escócia, que tinha o melhor curso de medicina da Grã-Bretanha. Darwin che-gou ali em 1825 e permaneceu no curso por dois anos.
Esse é um período muito importante, no qual ele se dedica intensamente a várias disciplinas, como química e geologia, aprende técnicas de taxidermia de aves, pesquisa os invertebrados marinhos e co-leta e classifica insetos – ou seja, estuda tudo, menos medicina. O trabalho com invertebrados marinhos tinha sido sugerido por Robert E. Grant (1793-1874), na época o maior especialista nesses animais da Grã-Bretanha. Médico formado
em Edimburgo, Grant dedicava-se à biologia e já havia publicado mais de 20 trabalhos científicos, alguns traduzidos na França. Era admirador incondicional tanto da Zoonomia do avô de Darwin quanto de Lamarck. Sob orientação de Grant, Darwin fez descobertas notáveis sobre a biologia dos briozoários (minúsculos animais que vivem presos a rochas e a outros seres). Os anos passados em Edimburgo deram ao jovem um sólido conhecimento de história natural, tanto prático quanto teórico – e essa teoria era lamarckista.
Ao perceber que o filho não tinha interesse pela medicina, Robert Darwin decide enviá-lo, aos 19 anos, para a Universidade de Cambridge, onde ele se prepararia para uma carreira de clérigo. Em Cambridge, ele conhece os clérigos naturalistas John Henslow (1796-1861) e Adam Sedgwick (1785-1873), com os quais estuda botânica e geologia. Nos quatro anos passados nessa universidade, continuou sua formação em ciências com excelentes cientis-tas, mas também homens religiosos que acreditavam em um Deus pessoal e, por outro lado, execravam teorias materialistas, como a teoria evolutiva de Lamarck.
Os dois anos em Edimburgo, du-rante os quais recebeu instrução em história natural, mas também uma doutrinação lamarckista, foram apa-gados nos quatro anos de Cambridge, no que se refere às teorias de La-marck. Quando Darwin iniciou sua longa viagem de circunavegação, em 1831, era um jovem de 22 anos que desejava ser um clérigo com uma paróquia no interior, cujos dí-zimos garantiriam sua manutenção e a de sua família, e onde poderia se dedicar com tranqüilidade a suas atividades científicas.
Erros grosseiros
Durante os cinco anos da viagem, ele coletou centenas de plantas e animais, desde zooplâncton até tartarugas e mamíferos fósseis, mas sem qualquer teoria evolutiva na cabeça. A ausência de uma teoria levou o jovem naturalista a cometer erros grosseiros. Ele comeu uma espécie nova de ema da Patagônia (na Argentina) e diferentes espécies de tartarugas das ilhas Galápagos (no oceano Pacífico), atirando os cascos pela borda do navio sem notar que em cada ilha existia uma forma diferente, e ainda coletou aves dessas mesmas ilhas sem anotar a ilha de origem de cada uma.
Por que tantos equívocos? Por que Darwin não percebeu a importância da flora e da fauna das ilhas Galápagos? A resposta é simples e está contida na famosa frase do ucraniano Theodosius Dobzhansky (1900-1975): “Nada em biologia faz sentido a não ser sob a luz da evolução.” E a teoria evolutiva só se-ria criada anos mais tarde pelo próprio Darwin. Esse é um excelente exemplo da função das teorias na capacidade de observar o mundo a nossa volta. Certos filósofos da ciência dizem que a observação só é possível quando o observador tem uma teoria que lhe permite fazer essa observação.
Primeiras contribuições
Após a viagem, Darwin instalou-se em Londres e redigiu seus pri-meiros trabalhos científicos sobre geologia. Estes fizeram enorme sucesso, o que é comprovado por sua eleição, em seguida, como se-cretário da Sociedade de Geologia. Nesse período começa a receber, do ornitólogo John Gould (1804-1881), informações sobre as aves das ilhas Galápagos. Gould informou que naquelas ilhas viviam 13 espécies novas, todas do mesmo grupo taxonômico e cada uma em sua ilha. São os pássaros conhecidos hoje como os fringilídeos de Darwin. Gould também descreveu a nova espécie de ema que o jovem na-turalista comeu, graças às penas e aos ossos que este guardou, após a refeição. O nome científico da ave, Rhea dar-winii, é uma homenagem de Gould ao amigo. Coube a Richard Owen (1804-1892), então o maior anatomista da Inglaterra, classificar os fósseis descobertos por Darwin, que revelaram a existência de uma preguiça e de uma lhama com tamanho muito superior ao das es-pécies atuais. Os mamíferos coletados na viagem foram classificados pelo zoólogo George Waterhouse (1810-1888), e Henslow fez o mesmo com as plantas.
Foram as informações desses taxonomistas que levaram Darwin, em 1838, a admitir a possibilidade da transmutação das espécies. Nesse mesmo ano ele leu o livro do primeiro economista inglês, Thomas R. Malthus (1766-1834), sobre a luta pela sobrevivência em um mundo onde a população cresce mais rápido que a produção de alimentos.
Darwin tinha conhecimento prático do crescimento da população: após seu casamento com a prima Emma, em 1839, ele teve nove filhos com intervalo médio de apenas 15 meses. Malthus afirmou que, se a população cresce mais que a produção de alimentos, é inevitável a competição desenfreada por recursos. Os agentes capazes de controlar o crescimento populacional, segundo o economista, são a fome, o vício e a miséria. Para ele, tais agentes foram criados por Deus como forma de acicate para estimular o desenvolvimento do homem, um ser naturalmente preguiçoso. Essa era a razão da posição de Malthus contra qualquer tipo de caridade. A caridade, dizia, atrapalharia o desenvolvimento natural do homem. São palavras estranhas na boca de um clérigo, mas não na boca de um economista com grandes contribuições no desenvolvimento do sistema econômico capitalista.
Com a idéia de que as espécies não são estáveis, de que elas se transformam com o passar do tempo – na verdade, uma idéia de Lamarck –, Darwin percebeu que estava se aliando a pessoas como Grant, aos socialistas que queriam eleições com voto universal, aos grupos que atacavam tanto a igreja da Inglaterra quanto o próprio sistema monárquico. Grant, ex-orientador de Darwin, trabalhava como professor da Universidade de Londres e havia sido expulso da So-ciedade Geológica por suas idéias lamarckistas; expulsão patrocina-da por Owen e Charles Lyell (1797-1875), este presidente da Socieda-de e grande amigo de Darwin. Lyell considerava uma estupidez a idéia de que as espécies se transformam. Se isso ocorre, dizia o geólogo, elas não existem, e isso é uma bobagem.
A teoria da evolução
Darwin vivia um enorme conflito, pois suas idéias o levavam a uma posição de enfrentamento em relação a todos os amigos cientistas, muitos clérigos anglicanos. Incapaz de enfrentar essa situação, ele esperou 20 anos para publicar, em 1859, seu livro mais importante, A origem das espécies, no qual formula a teoria da evolução baseada no surgimento de variabilidade genética ao acaso e da ação da seleção natural sobre essa variabilidade. Nenhum aspecto essencial dessa teoria foi alterado nos últimos 150 anos, o que é um fato notável.
A qualidade da teoria darwi-nista deve-se em parte à excelên-cia atingida por Darwin na área da taxonomia. Nos anos 1840, quan-do tentava convencer o botânico Joseph D. Hooker (1817-1911), dire-tor do Jardim Botânico Real, em Kew, a aceitar suas idéias, recebeu uma carta do amigo na qual este se queixava de alguns biólogos que opinavam sobre espécies sem nunca ter descrito uma. Darwin ‘vestiu a carapuça’, já que se enquadra-va nesse perfil, e decidiu sanar a falha. Durante sete anos dedicou-se a descrever as espécies de cracas, atuais e fósseis, publicando, sobre a taxonomia desses animais, várias monografias que são uma refe-rência útil ainda hoje. É um trabalho monumental que inscreveu Darwin entre os melhores taxonomistas da época.
O conhecimento obtido com a taxonomia das cracas forneceu ao cientista uma percepção profunda dos mecanismos que geram novas espécies – as espécies em estado nascendi – e permitiu a ele inven-tar não apenas a teoria da evolução, o que já seria um grande trabalho, mas também uma nova disciplina, a biogeografia. Darwin publicou mais de uma dezena de livros importantes, sendo o mais famoso A origem das espécies, que bem poucas pessoas, incluindo muitos biólogos, leram na íntegra. É uma pena, pois essa obra, com quase 150 anos de idade, ainda é muito atual.
O fato de muitos biólogos importantes na atualidade não terem lido A origem das espécies pode explicar por que ele está ausente das citações bibliográficas em livros recentes e por que o cientista inglês não é citado como autor de conceitos importantes, como ni-cho ecológico, ecossistemas, separação de caracteres em função da competição, variabilidade genéti-ca surgida ao acaso e muitos outros. Tais conceitos são atribuídos a cien-tistas modernos, provavelmente porque não leram atentamente o livro de Darwin. Mas ainda há tem-po. Ler A origem das espécies com atenção seria um conselho importante até para famosos neodar-winistas radicais, como o biólogo britânico Richard Dawkins, pois nesse livro Darwin nos diz que na evolução das espécies nem tudo pode ser explicado pela seleção natural!
Por
Ricardo Iglesias Rios
Departamento de Ecologia, Instituto de Biologia,
Universidade Federal do Rio de Janeiro
CIÊNCIA HOJE • vol. 42 • nº 250
Nascido no século 19, Charles Robert Darwin (1809-1882) é o cientista mais conhecido e reconhecido no século 21, superando em notoriedade gigantes como o físico inglês Isaac Newton (1643-1727), o filósofo alemão Karl Marx (1818-1883), o neurologista austría-co Sigmund Freud (1856-1939) e o físico alemão Albert Einstein (1879-1955). Darwin foi o criador da teoria evolutiva que explica a origem de todas as espécies por meio da seleção natural. Essa teoria, após ter sido elaborada por ele durante décadas, foi lançada a público há exatos 150 anos, com a leitura, em sessão da Royal Society, principal academia de ciências da Inglater-ra, de cartas do próprio Darwin e do também inglês Alfred Wallace (1823-1913), que chegou bem mais tarde a conclusões semelhantes. Embora tenha mudado radicalmente a forma como o homem percebia a natureza, a teoria da evolução dificilmente explica o enorme prestígio que Darwin tem ainda hoje.
Na verdade, a contribuição de Darwin para o desenvolvimento do conhecimento humano vai muito além da teoria evolutiva: ele foi pioneiro na geologia, criando teorias importantes para o surgimento de ilhas oceânicas e explicações corretas para os mecanismos que geram os movimentos ascendentes da cordilheira dos Andes. Para o biólogo austríaco Konrad Lorenz (1903-1989), Darwin foi ainda o iniciador da etologia, ciência que estuda o comportamento dos animais. Além disso, pode ser considerado o mentor da moderna ecologia, por ter criado os conceitos de ‘nicho ecológico’ e ‘ecossistema’, ao afirmar que cada espécie ocupa um lugar determinado (nicho) na “economia da natureza” (ecossistema). Nessa mesma área, Darwin demonstrou a importância funda-mental, para o funcionamento dos ecossistemas, de relações como competição, predação e mutualismo entre os seres vivos e interações entre estes e o ambiente. Na taxonomia, a rainha das ciências biológicas, ele fez contribuições notáveis, criando os conceitos de ‘espécies em estado nascendi’ e de ‘espécie fóssil’. Naturalista, biólogo, etólogo, taxonomista, geólogo, ecó-logo e, como se fosse pouco, pai da biogeografia. Suas contribuições fundamentais em todas essas ciências podem explicar por que Dar-win é um gigante entre gigantes.
De avô para neto
O criador da teoria da evolução nasceu em 1809 em uma família rica, com tradição na medicina. O pai, Robert Darwin (1766-1848), era médico, e o avô paterno, Erasmus Darwin (1731-1802) era, além de médico, um poeta e um adepto das idéias sobre evolução do biólogo francês Jean-Baptiste Lamarck (1744-1829), que acreditava na transmutação das espécies e criou uma teoria para explicá-la. Erasmus escreveu Zoonomia, um vo-lumoso livro sobre o tema da ori-gem das espécies, lido e apreciado pelo neto. Aos oito anos, Charles Darwin perdeu a mãe e entrou pa-ra a escola. Os oito anos passados nessa escola foram considerados pelo próprio Darwin uma grande e enfadonha perda de tempo. Seu baixo desempenho fez com que seu pai o enviasse para a Universidade de Edimburgo, na Escócia, que tinha o melhor curso de medicina da Grã-Bretanha. Darwin che-gou ali em 1825 e permaneceu no curso por dois anos.
Esse é um período muito importante, no qual ele se dedica intensamente a várias disciplinas, como química e geologia, aprende técnicas de taxidermia de aves, pesquisa os invertebrados marinhos e co-leta e classifica insetos – ou seja, estuda tudo, menos medicina. O trabalho com invertebrados marinhos tinha sido sugerido por Robert E. Grant (1793-1874), na época o maior especialista nesses animais da Grã-Bretanha. Médico formado
em Edimburgo, Grant dedicava-se à biologia e já havia publicado mais de 20 trabalhos científicos, alguns traduzidos na França. Era admirador incondicional tanto da Zoonomia do avô de Darwin quanto de Lamarck. Sob orientação de Grant, Darwin fez descobertas notáveis sobre a biologia dos briozoários (minúsculos animais que vivem presos a rochas e a outros seres). Os anos passados em Edimburgo deram ao jovem um sólido conhecimento de história natural, tanto prático quanto teórico – e essa teoria era lamarckista.
Ao perceber que o filho não tinha interesse pela medicina, Robert Darwin decide enviá-lo, aos 19 anos, para a Universidade de Cambridge, onde ele se prepararia para uma carreira de clérigo. Em Cambridge, ele conhece os clérigos naturalistas John Henslow (1796-1861) e Adam Sedgwick (1785-1873), com os quais estuda botânica e geologia. Nos quatro anos passados nessa universidade, continuou sua formação em ciências com excelentes cientis-tas, mas também homens religiosos que acreditavam em um Deus pessoal e, por outro lado, execravam teorias materialistas, como a teoria evolutiva de Lamarck.
Os dois anos em Edimburgo, du-rante os quais recebeu instrução em história natural, mas também uma doutrinação lamarckista, foram apa-gados nos quatro anos de Cambridge, no que se refere às teorias de La-marck. Quando Darwin iniciou sua longa viagem de circunavegação, em 1831, era um jovem de 22 anos que desejava ser um clérigo com uma paróquia no interior, cujos dí-zimos garantiriam sua manutenção e a de sua família, e onde poderia se dedicar com tranqüilidade a suas atividades científicas.
Erros grosseiros
Durante os cinco anos da viagem, ele coletou centenas de plantas e animais, desde zooplâncton até tartarugas e mamíferos fósseis, mas sem qualquer teoria evolutiva na cabeça. A ausência de uma teoria levou o jovem naturalista a cometer erros grosseiros. Ele comeu uma espécie nova de ema da Patagônia (na Argentina) e diferentes espécies de tartarugas das ilhas Galápagos (no oceano Pacífico), atirando os cascos pela borda do navio sem notar que em cada ilha existia uma forma diferente, e ainda coletou aves dessas mesmas ilhas sem anotar a ilha de origem de cada uma.
Por que tantos equívocos? Por que Darwin não percebeu a importância da flora e da fauna das ilhas Galápagos? A resposta é simples e está contida na famosa frase do ucraniano Theodosius Dobzhansky (1900-1975): “Nada em biologia faz sentido a não ser sob a luz da evolução.” E a teoria evolutiva só se-ria criada anos mais tarde pelo próprio Darwin. Esse é um excelente exemplo da função das teorias na capacidade de observar o mundo a nossa volta. Certos filósofos da ciência dizem que a observação só é possível quando o observador tem uma teoria que lhe permite fazer essa observação.
Primeiras contribuições
Após a viagem, Darwin instalou-se em Londres e redigiu seus pri-meiros trabalhos científicos sobre geologia. Estes fizeram enorme sucesso, o que é comprovado por sua eleição, em seguida, como se-cretário da Sociedade de Geologia. Nesse período começa a receber, do ornitólogo John Gould (1804-1881), informações sobre as aves das ilhas Galápagos. Gould informou que naquelas ilhas viviam 13 espécies novas, todas do mesmo grupo taxonômico e cada uma em sua ilha. São os pássaros conhecidos hoje como os fringilídeos de Darwin. Gould também descreveu a nova espécie de ema que o jovem na-turalista comeu, graças às penas e aos ossos que este guardou, após a refeição. O nome científico da ave, Rhea dar-winii, é uma homenagem de Gould ao amigo. Coube a Richard Owen (1804-1892), então o maior anatomista da Inglaterra, classificar os fósseis descobertos por Darwin, que revelaram a existência de uma preguiça e de uma lhama com tamanho muito superior ao das es-pécies atuais. Os mamíferos coletados na viagem foram classificados pelo zoólogo George Waterhouse (1810-1888), e Henslow fez o mesmo com as plantas.
Foram as informações desses taxonomistas que levaram Darwin, em 1838, a admitir a possibilidade da transmutação das espécies. Nesse mesmo ano ele leu o livro do primeiro economista inglês, Thomas R. Malthus (1766-1834), sobre a luta pela sobrevivência em um mundo onde a população cresce mais rápido que a produção de alimentos.
Darwin tinha conhecimento prático do crescimento da população: após seu casamento com a prima Emma, em 1839, ele teve nove filhos com intervalo médio de apenas 15 meses. Malthus afirmou que, se a população cresce mais que a produção de alimentos, é inevitável a competição desenfreada por recursos. Os agentes capazes de controlar o crescimento populacional, segundo o economista, são a fome, o vício e a miséria. Para ele, tais agentes foram criados por Deus como forma de acicate para estimular o desenvolvimento do homem, um ser naturalmente preguiçoso. Essa era a razão da posição de Malthus contra qualquer tipo de caridade. A caridade, dizia, atrapalharia o desenvolvimento natural do homem. São palavras estranhas na boca de um clérigo, mas não na boca de um economista com grandes contribuições no desenvolvimento do sistema econômico capitalista.
Com a idéia de que as espécies não são estáveis, de que elas se transformam com o passar do tempo – na verdade, uma idéia de Lamarck –, Darwin percebeu que estava se aliando a pessoas como Grant, aos socialistas que queriam eleições com voto universal, aos grupos que atacavam tanto a igreja da Inglaterra quanto o próprio sistema monárquico. Grant, ex-orientador de Darwin, trabalhava como professor da Universidade de Londres e havia sido expulso da So-ciedade Geológica por suas idéias lamarckistas; expulsão patrocina-da por Owen e Charles Lyell (1797-1875), este presidente da Socieda-de e grande amigo de Darwin. Lyell considerava uma estupidez a idéia de que as espécies se transformam. Se isso ocorre, dizia o geólogo, elas não existem, e isso é uma bobagem.
A teoria da evolução
Darwin vivia um enorme conflito, pois suas idéias o levavam a uma posição de enfrentamento em relação a todos os amigos cientistas, muitos clérigos anglicanos. Incapaz de enfrentar essa situação, ele esperou 20 anos para publicar, em 1859, seu livro mais importante, A origem das espécies, no qual formula a teoria da evolução baseada no surgimento de variabilidade genética ao acaso e da ação da seleção natural sobre essa variabilidade. Nenhum aspecto essencial dessa teoria foi alterado nos últimos 150 anos, o que é um fato notável.
A qualidade da teoria darwi-nista deve-se em parte à excelên-cia atingida por Darwin na área da taxonomia. Nos anos 1840, quan-do tentava convencer o botânico Joseph D. Hooker (1817-1911), dire-tor do Jardim Botânico Real, em Kew, a aceitar suas idéias, recebeu uma carta do amigo na qual este se queixava de alguns biólogos que opinavam sobre espécies sem nunca ter descrito uma. Darwin ‘vestiu a carapuça’, já que se enquadra-va nesse perfil, e decidiu sanar a falha. Durante sete anos dedicou-se a descrever as espécies de cracas, atuais e fósseis, publicando, sobre a taxonomia desses animais, várias monografias que são uma refe-rência útil ainda hoje. É um trabalho monumental que inscreveu Darwin entre os melhores taxonomistas da época.
O conhecimento obtido com a taxonomia das cracas forneceu ao cientista uma percepção profunda dos mecanismos que geram novas espécies – as espécies em estado nascendi – e permitiu a ele inven-tar não apenas a teoria da evolução, o que já seria um grande trabalho, mas também uma nova disciplina, a biogeografia. Darwin publicou mais de uma dezena de livros importantes, sendo o mais famoso A origem das espécies, que bem poucas pessoas, incluindo muitos biólogos, leram na íntegra. É uma pena, pois essa obra, com quase 150 anos de idade, ainda é muito atual.
O fato de muitos biólogos importantes na atualidade não terem lido A origem das espécies pode explicar por que ele está ausente das citações bibliográficas em livros recentes e por que o cientista inglês não é citado como autor de conceitos importantes, como ni-cho ecológico, ecossistemas, separação de caracteres em função da competição, variabilidade genéti-ca surgida ao acaso e muitos outros. Tais conceitos são atribuídos a cien-tistas modernos, provavelmente porque não leram atentamente o livro de Darwin. Mas ainda há tem-po. Ler A origem das espécies com atenção seria um conselho importante até para famosos neodar-winistas radicais, como o biólogo britânico Richard Dawkins, pois nesse livro Darwin nos diz que na evolução das espécies nem tudo pode ser explicado pela seleção natural!
Por
Ricardo Iglesias Rios
Departamento de Ecologia, Instituto de Biologia,
Universidade Federal do Rio de Janeiro
CIÊNCIA HOJE • vol. 42 • nº 250

Dinosaurs take to the air
Flying birds evolved from a group of bipedal dinosaurs. The latest fossil discoveries from China indicate that the dinosaurian ancestors of birds had four wings — and that these animals glided rather than flapped.
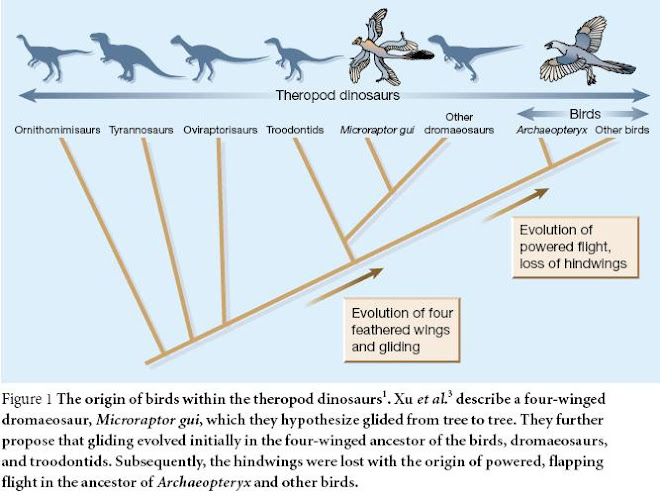
Three questions lie at the heart of the debate about the evolution of birds: theorigin of the group itself, the origin of feathers and the origin of flight. Progress in reconstructing the evolutionary history of dinosaurs has established a well-corroborated answer to the first question1. Birds are a lineage of dinosaurs, and are most closely related to dromaeosaurs and troodontids, both of which belong to a group of bipedal, carnivorous dinosaurs called theropods. Since 1997, new fossils and insights from developmental biology have also supported a coherent solution to the second question2. Feathers evolved in theropod dinosaurs before the origin of birds or flight through a series of developmental novelties. Now, Xu et al.3 report spectacular 124–128-million-year-old fossils from Liaoning, China, that promise to revolutionize discussion of the last question — how did avian flight evolve? For more than a century, debate on the origin of bird flight has centred on two different hypotheses4. According to arboreal theories, flight arose in tree-dwelling creatures through an intermediate gliding stage, an idea that has been supported by the observation that flight is energetically more efficient at higher speeds (when more lift is generated)5. Further, the flight stroke in continuous, level flight is simpler than in take-off from the ground. According to the competing, cursorial theories, flight evolved in ground-living animals via a powered running stage. This view is supported by the evidence that birds evolved from a lineage of terrestrial, bipedal theropods, and that many components of the avian flight apparatus evolved originally in a terrestrial context4. Moreover, aerodynamic models6 of the flight stroke of Archaeopteryx, the earliest bird accepted as such, indicate that its wings could have provided thrust as well as lift, and aided the legs in achieving enough ground speed for a running take-off. In a colourful and prescient paper of 1915, however, William Beebe7 proposed that avian flight evolved through a gliding, four-winged — tetrapteryx — stage with wing feathers on both the arms and the legs. Now Xu and colleagues3 describe a small dromaeosaur, Microraptor gui, that sports four wings of fully modern, asymmetrical feathers on its forelimbs and legs, and looks as if it could have glided straight out of the pages of Beebe’s notebooks. Although the specimens of Microraptor are younger than those of Archaeopteryx, Microraptor is a basal member — an early evolutionary branch — of the closest relatives of Archaeopteryx and other birds (Fig. 1). In support of the arboreal theory, Xu et al. propose that the most recent common ancestor of birds and dromaeosaurs was a four-winged creature that lived in and glided among trees. The evidence of an aerodynamic function for Microraptor’s forelimb and leg feathers is excellent. Asymmetrical feather vanes have long been recognized as indicating aerodynamic function in flight or gliding. Furthermore, the feathers of both the forewings and the hindwings increase in asymmetry towards the end of the limb in a striking match with the primary and secondary feathers of a modern bird wing. The discovery of dromaeosaur hindwings was presaged last year by the description of 13- cm-long modern, vaned feathers on the tibia of an even larger, unnamed dromaeosaur from Liaoning8, but that specimen lacked enough detail to describe the entire structure. The long tail of Microraptor also features a terminal tuft of feathers like that found on other basal dromaeosaurs8 and on another theropod, the oviraptoran Caudipteryx9. The discovery of a logical functional intermediate provides striking support for the arboreal–gliding hypothesis of the origin of bird flight. However, substantial questions remain. In particular, how did Microraptor actually use its four wings? Perhaps because flapping hindwings are so unlikely, Xu et al. conclude that Microraptor merely glided, and did not have a powered flight stroke. Palaeontologists and functional morphologists will be eager to study the shoulder and wing anatomy to judge whether Microraptor could sustain powered flight. More information is also required on how the animal could have rotated its legs to deploy its hindwings. Xu et al. maintain that all four wings were present in the earliest ancestors of birds and dromaeosaurs, and that with the evolution of powered flight the hindwings were lost in the avian lineage before the advent of Archaeopteryx. It is also possible, however, that the hindwings are a unique feature of dromaeosaurs. Palaeontologists will want to re-examine specimens of Archaeopteryx for any evidence of a vestigial hindwing (Beebe found none). Regardless of the upshot of those enquiries, there is no doubt that the forewings of Microraptor, Archaeopteryx and other birds are homologous and had an aerodynamic function. Xu et al. also argue that the extensively feathered legs of Microraptorwould have been incompatible with life on the ground. The feathers extend all the way down the leg, much further than they do in Beebe’s mythical tetrapteryx. Dragging your wing feathers in the dirt would doubtless be aerodynamically disadvantageous, but it will require detailed reconstructions of Microraptor’s hindlimbs with feathers attached to rule out the possibility that it could have walked and run. Finally, although Xu and colleagues report that Microraptor has the anatomical features of a dromaeosaur, firm conclusions about the evolution of bird flight will require new systematic analyses incorporating this and other newly discovered theropod species from Liaoning to confirm their phylogenetic position. Sceptics will argue in any case that Microraptor and dromaeosaurs are more closely related to modern birds than is Archaeopteryx — but then they will also have to address the problem of why a bird that could flap its wings perfectly well would evolve a second pair of wings. Birds are traditionally considered to be animals with a difference: that is, to be a distinct vertebrate class despite their origins within the reptiles. But advances in palaeontology, phylogenetics and evolutionary biology have erased the anatomical gap between birds and their dinosaur ancestors10. Now that dromaeosaurs have taken to the air, in the form of Microraptor, there remain no major traits that are unique to birds — with the possible exception of powered flight. Although some may be irked at this lost distinction, the benefits will be a fuller, more integrated understanding of avian biology. This new evidence of an arboreal, gliding stage in the evolution of bird flight complements the evidence of terrestrial evolution in the theropod dinosaurs. Terrestrial theropod dinosaurs had evolved for millions of years before the ancestors of Microraptor and the birds took to the trees or to the air. Moving beyond the arboreal versus cursorial debate over the origins of bird flight4, the task ahead is to understand which components of the avian flight apparatus evolved in a terrestrial and which in an arboreal context.
Nature 2008
Richard O. Prum is in the Department of Ecology
and Evolutionary Biology, and the Natural History
Museum and Biodiversity Research Center,
University of Kansas, Lawrence, Kansas 66045, USA.
e-mail: prum@ku.edu
1. Sereno, P. Science 284, 2137–2147 (1999).
2. Prum, R. O. & Brush, A. H. Q. Rev. Biol. 77, 261–295 (2002).
3. Xu, X. et al. Nature 421, 335–340 (2003).
4. Padian, K. & Chiappe, L. M. Biol. Rev. 73, 1–42 (1998).
5. Rayner, J. M. V. in The Beginnings of Birds (eds Hecht, M. K.,
Ostrom, J. H., Viohl, G. & Wellnhofer, P.) 279–287 (Freunde
des Jura-Museum, Eichstätt, 1985).
6. Burgers, P. & Chiappe, L. M. Nature 399, 60–62 (1999).
7. Beebe, W. H. Zoologica 2, 38–52 (1915).
8. Norell, M. et al. Nature 416, 36–37 (2002).
9. Ji, Q., Currie, P. J., Norell, M. A. & Ji, S.-A. Nature 393, 753–761 (1998).
10.Padian, K. Nature 393, 729–730 (1998).
Three questions lie at the heart of the debate about the evolution of birds: theorigin of the group itself, the origin of feathers and the origin of flight. Progress in reconstructing the evolutionary history of dinosaurs has established a well-corroborated answer to the first question1. Birds are a lineage of dinosaurs, and are most closely related to dromaeosaurs and troodontids, both of which belong to a group of bipedal, carnivorous dinosaurs called theropods. Since 1997, new fossils and insights from developmental biology have also supported a coherent solution to the second question2. Feathers evolved in theropod dinosaurs before the origin of birds or flight through a series of developmental novelties. Now, Xu et al.3 report spectacular 124–128-million-year-old fossils from Liaoning, China, that promise to revolutionize discussion of the last question — how did avian flight evolve? For more than a century, debate on the origin of bird flight has centred on two different hypotheses4. According to arboreal theories, flight arose in tree-dwelling creatures through an intermediate gliding stage, an idea that has been supported by the observation that flight is energetically more efficient at higher speeds (when more lift is generated)5. Further, the flight stroke in continuous, level flight is simpler than in take-off from the ground. According to the competing, cursorial theories, flight evolved in ground-living animals via a powered running stage. This view is supported by the evidence that birds evolved from a lineage of terrestrial, bipedal theropods, and that many components of the avian flight apparatus evolved originally in a terrestrial context4. Moreover, aerodynamic models6 of the flight stroke of Archaeopteryx, the earliest bird accepted as such, indicate that its wings could have provided thrust as well as lift, and aided the legs in achieving enough ground speed for a running take-off. In a colourful and prescient paper of 1915, however, William Beebe7 proposed that avian flight evolved through a gliding, four-winged — tetrapteryx — stage with wing feathers on both the arms and the legs. Now Xu and colleagues3 describe a small dromaeosaur, Microraptor gui, that sports four wings of fully modern, asymmetrical feathers on its forelimbs and legs, and looks as if it could have glided straight out of the pages of Beebe’s notebooks. Although the specimens of Microraptor are younger than those of Archaeopteryx, Microraptor is a basal member — an early evolutionary branch — of the closest relatives of Archaeopteryx and other birds (Fig. 1). In support of the arboreal theory, Xu et al. propose that the most recent common ancestor of birds and dromaeosaurs was a four-winged creature that lived in and glided among trees. The evidence of an aerodynamic function for Microraptor’s forelimb and leg feathers is excellent. Asymmetrical feather vanes have long been recognized as indicating aerodynamic function in flight or gliding. Furthermore, the feathers of both the forewings and the hindwings increase in asymmetry towards the end of the limb in a striking match with the primary and secondary feathers of a modern bird wing. The discovery of dromaeosaur hindwings was presaged last year by the description of 13- cm-long modern, vaned feathers on the tibia of an even larger, unnamed dromaeosaur from Liaoning8, but that specimen lacked enough detail to describe the entire structure. The long tail of Microraptor also features a terminal tuft of feathers like that found on other basal dromaeosaurs8 and on another theropod, the oviraptoran Caudipteryx9. The discovery of a logical functional intermediate provides striking support for the arboreal–gliding hypothesis of the origin of bird flight. However, substantial questions remain. In particular, how did Microraptor actually use its four wings? Perhaps because flapping hindwings are so unlikely, Xu et al. conclude that Microraptor merely glided, and did not have a powered flight stroke. Palaeontologists and functional morphologists will be eager to study the shoulder and wing anatomy to judge whether Microraptor could sustain powered flight. More information is also required on how the animal could have rotated its legs to deploy its hindwings. Xu et al. maintain that all four wings were present in the earliest ancestors of birds and dromaeosaurs, and that with the evolution of powered flight the hindwings were lost in the avian lineage before the advent of Archaeopteryx. It is also possible, however, that the hindwings are a unique feature of dromaeosaurs. Palaeontologists will want to re-examine specimens of Archaeopteryx for any evidence of a vestigial hindwing (Beebe found none). Regardless of the upshot of those enquiries, there is no doubt that the forewings of Microraptor, Archaeopteryx and other birds are homologous and had an aerodynamic function. Xu et al. also argue that the extensively feathered legs of Microraptorwould have been incompatible with life on the ground. The feathers extend all the way down the leg, much further than they do in Beebe’s mythical tetrapteryx. Dragging your wing feathers in the dirt would doubtless be aerodynamically disadvantageous, but it will require detailed reconstructions of Microraptor’s hindlimbs with feathers attached to rule out the possibility that it could have walked and run. Finally, although Xu and colleagues report that Microraptor has the anatomical features of a dromaeosaur, firm conclusions about the evolution of bird flight will require new systematic analyses incorporating this and other newly discovered theropod species from Liaoning to confirm their phylogenetic position. Sceptics will argue in any case that Microraptor and dromaeosaurs are more closely related to modern birds than is Archaeopteryx — but then they will also have to address the problem of why a bird that could flap its wings perfectly well would evolve a second pair of wings. Birds are traditionally considered to be animals with a difference: that is, to be a distinct vertebrate class despite their origins within the reptiles. But advances in palaeontology, phylogenetics and evolutionary biology have erased the anatomical gap between birds and their dinosaur ancestors10. Now that dromaeosaurs have taken to the air, in the form of Microraptor, there remain no major traits that are unique to birds — with the possible exception of powered flight. Although some may be irked at this lost distinction, the benefits will be a fuller, more integrated understanding of avian biology. This new evidence of an arboreal, gliding stage in the evolution of bird flight complements the evidence of terrestrial evolution in the theropod dinosaurs. Terrestrial theropod dinosaurs had evolved for millions of years before the ancestors of Microraptor and the birds took to the trees or to the air. Moving beyond the arboreal versus cursorial debate over the origins of bird flight4, the task ahead is to understand which components of the avian flight apparatus evolved in a terrestrial and which in an arboreal context.
Nature 2008
Richard O. Prum is in the Department of Ecology
and Evolutionary Biology, and the Natural History
Museum and Biodiversity Research Center,
University of Kansas, Lawrence, Kansas 66045, USA.
e-mail: prum@ku.edu
1. Sereno, P. Science 284, 2137–2147 (1999).
2. Prum, R. O. & Brush, A. H. Q. Rev. Biol. 77, 261–295 (2002).
3. Xu, X. et al. Nature 421, 335–340 (2003).
4. Padian, K. & Chiappe, L. M. Biol. Rev. 73, 1–42 (1998).
5. Rayner, J. M. V. in The Beginnings of Birds (eds Hecht, M. K.,
Ostrom, J. H., Viohl, G. & Wellnhofer, P.) 279–287 (Freunde
des Jura-Museum, Eichstätt, 1985).
6. Burgers, P. & Chiappe, L. M. Nature 399, 60–62 (1999).
7. Beebe, W. H. Zoologica 2, 38–52 (1915).
8. Norell, M. et al. Nature 416, 36–37 (2002).
9. Ji, Q., Currie, P. J., Norell, M. A. & Ji, S.-A. Nature 393, 753–761 (1998).
10.Padian, K. Nature 393, 729–730 (1998).
Dinosaurs take to the air

Biomechanics: Independent evolution of running in vampire bats
Most tetrapods have retained terrestrial locomotion since it evolved in the Palaeozoic era1,2, but bats have become so specialized for flight that they have almost lost the ability to manoeuvre on land at all3,4. Vampire bats, which sneakup on their prey along the ground, are an important exception. Here we show that common vampire bats can also run byusing a unique bounding gait, in which the forelimbs instead of the hindlimbs are recruited for force production as the wingsare much more powerful than the legs. This ability to run seems to have evolved independently within the bat lineage.Bats (Chiroptera) are the only mammals that fly, so their bodies differ from those of terrestrial mammals. As a result, most grounded bats can only shuffle awkwardly rom a sprawled position4. However, the common vampire bat (Desmodus rotundus) walks forwards, sideways and backwards5, and initiates flight with a single vertical jumpfrom standing6. Captive D. rotundus have also been found to ‘hop’ at speeds exceeding 2.0 metres per second5.To determine whether this hopping behaviour constitutes a stereotyped running gait by D. rotundus, we tested five adultmales on a treadmill inside a Plexiglas cage. The animals used a walking gait at low treadmill speeds (0.12 to 0.56 m s_1) anda stereotyped running gait at high speeds (0.28 to 1.14 m s_1). The walking gait was similar to the typical lateral-sequence walking gait of other tetrapods7; however, therun was different from any gait previously described.We classify this novel gait as a run because it includes anotable aerial phase. A tetrapod typically increases its speed while walking by increasing its stride frequency. Atsome transition speed, animals switch to a running gait that permitsa further increase in speed, but at stride frequenciesthat are lower than would be predicted for high-speed walking8,9. Our kinematic data from D. rotundus fit this general stridefrequency– velocity relationship. , theslopes of the stride-frequency– velocity regressions,which are best fits to the walking and running data, respectively, and are shown truncated at the intersection, aresignificantly different (t-test, P_0.0001, n_61). These regression lines indicate that common vampire bats, like other running tetrapods, keep their stride frequencies low by walking at low speeds and running at high speeds .The walking vampire bats used stride frequencies that were comparable to those of similarly sized terrestrial mammals (mice) over the same range of speeds. When running, however, the bats used lower stride frequencies than mice8: this could be explained by the vampire bats’ longforearms, which allow longer and fewer strides to be taken during running than can be achieved by mice.The absence of a running gait in all other bat species so far surveyed indicates that running may have been lost early in the evolution of bats, evolving afresh in the vampires at a later time. We have shown that the hopping behaviour reported forD.rotundus in captivity5 is a running gait.But despite detailed knowledge of their roosting and foraging behaviour10,11, the selective benefit of running for these bats in the wild is not known.Presumably, vampire bats are most likely to run when manoeuvring around prey animals while feeding, and they may have used the gait more before the introduction of domestic livestock to the Americas in the sixteenth century11.
Daniel K. Riskin, John W.Hermanson
Department of Biomedical Sciences, College ofVeterinary Medicine, Cornell University, Ithaca,New York 14853, USAe-mail: dkr8@cornell.edu
1. Gambaryan, P. P. Zh. Obshch. Biol. 63, 426–445 (2002).
2. Parchman, A. J., Reilly, S. M. & Biknevicius, A. R. J. Exp. Biol.206, 1379–1388 (2003).
3. Vaughan, T. A. Univ. Kansas Publ.Mus. Nat. Hist. 12, 1–153(1959).
4. Riskin, D. K., Bertram, J. E. A. & Hermanson, J.W. J. Exp. Biol. (in the press).
5. Altenbach, J. S. Spec. Publ. Am. Soc.Mammal. 6, 137 (1979). 6. Schutt,W. A. Jr et al. J. Exp. Biol. 200, 3003–3012 (1997).7. Hildebrand, M. in Functional Vertebrate Morphology (edsHildebrand, M., Bramble, D. M., Liem, K. F. & Wake, D. B.)38–57 (Belknap, Harvard Univ. Press, Cambridge, 1985).8. Heglund, N. & Taylor, C. J. Exp. Biol. 138, 301–318 (1988).9. Taylor, C., Heglund, N. & Maloiy, G. J. Exp. Biol. 97, 1–21(1982).10.Turner, D. C. The Vampire Bat: A Field Study in Behavior and Ecology (Johns Hopkins Univ. Press, Baltimore, 1975).11.Greenhall, A. M. & Schmidt, U. Natural History of Vampire Bats (CRC, Boca Raton, 1988).
Supplementary information accompanies the paper on Nature’s website.
Competing financial interests: declared none.brief communications is intended to provide a forumfor short, topical reports of general scientific interest andfor online technical discussion of recently publishedmaterial of particular interest to non-specialist readers(brief communications arising). Priority will be givento contributions that have fewer than 500 words, 10references and one figure. Detailed guidelines areavailable on Nature’s website.
Daniel K. Riskin, John W.Hermanson
Department of Biomedical Sciences, College ofVeterinary Medicine, Cornell University, Ithaca,New York 14853, USAe-mail: dkr8@cornell.edu
1. Gambaryan, P. P. Zh. Obshch. Biol. 63, 426–445 (2002).
2. Parchman, A. J., Reilly, S. M. & Biknevicius, A. R. J. Exp. Biol.206, 1379–1388 (2003).
3. Vaughan, T. A. Univ. Kansas Publ.Mus. Nat. Hist. 12, 1–153(1959).
4. Riskin, D. K., Bertram, J. E. A. & Hermanson, J.W. J. Exp. Biol. (in the press).
5. Altenbach, J. S. Spec. Publ. Am. Soc.Mammal. 6, 137 (1979). 6. Schutt,W. A. Jr et al. J. Exp. Biol. 200, 3003–3012 (1997).7. Hildebrand, M. in Functional Vertebrate Morphology (edsHildebrand, M., Bramble, D. M., Liem, K. F. & Wake, D. B.)38–57 (Belknap, Harvard Univ. Press, Cambridge, 1985).8. Heglund, N. & Taylor, C. J. Exp. Biol. 138, 301–318 (1988).9. Taylor, C., Heglund, N. & Maloiy, G. J. Exp. Biol. 97, 1–21(1982).10.Turner, D. C. The Vampire Bat: A Field Study in Behavior and Ecology (Johns Hopkins Univ. Press, Baltimore, 1975).11.Greenhall, A. M. & Schmidt, U. Natural History of Vampire Bats (CRC, Boca Raton, 1988).
Supplementary information accompanies the paper on Nature’s website.
Competing financial interests: declared none.brief communications is intended to provide a forumfor short, topical reports of general scientific interest andfor online technical discussion of recently publishedmaterial of particular interest to non-specialist readers(brief communications arising). Priority will be givento contributions that have fewer than 500 words, 10references and one figure. Detailed guidelines areavailable on Nature’s website.
Biomechanics: Independent evolution of running in vampire bats Fig1

Biomechanics: Independent evolution of running in vampire bats Fig 2.



















































